概要
m6A、m1A、m5C、およびシュードウリジンなどのRNA修飾は、エピトランスクリプトーム、および遺伝子発現制御に深く関係しています。特にm6Aは、mRNAおよびlncRNAにおけるもっとも豊富な内部修飾となっており、転写後mRNA/lncRNA代謝、および機能のあらゆる局面に影響を与えます。さらに、m6Aは、キャップに依存しないcircRNAの翻訳開始やPrimary microRNA(pri-miRNA)のプロセシングなど、他の多くのncRNAの機能にも関与しています。
RNAの潜在的な影響は、遺伝子転写物だけでなく、転写物の修飾率にも影響します。しかしながら、現在のトランスクリプトームワイドのRNA修飾プロファイリングにおいては、主に修飾部位のマッピングは扱うものの、その転写物についての修飾RNAの割合を定量化するところまでは至っていませんでした。
特長
本サービスで使用するエピトランスクリプトームマイクロアレイは、2色チャンネルマイクロアレイテクノロジーと、RNA修飾免疫沈降法を組み合わせて、転写産物をアイソフォーム特異的にRNAの修飾レベルの割合を定量化します。このマイクロアレイはmRNA、lncRNA、circRNA、pre-miRNA、pri-miRNA、snoRNA、およびsnRNAの各クラスのエピトランスクリプトームをカバーしています。
エピトランスクリプトームマイクロアレイは、シークエンスによるRNAメチル化解析(MeRIP-Seq)よりも優れた利点を持っています。
- どの遺伝子転写産物が修飾されているか、条件間の差動的な修飾、各転写産物に対する修飾の割合を単一のマイクロアレイで同時にプロファイルすることが可能です。
- MeRIP-Seqでプロファイリングが困難なlncRNAやcircRNAであっても、コード、ノンコードのRNAクラスのカバレッジが可能です。
- rRNAの除去が不要で、MeRIPよりも迅速かつ簡単に解析が行えます。
- 少量サンプルでの解析が可能で、3μgのtotalRNAから解析が可能です。
*フィルジェン社では海外輸送ならびにサンプルQC使用のため、10μg以上をご準備頂くことを推奨しております。
【少量サンプルでの解析】
現在のMeRIP-Seq法では、スタートサンプルとして120μg以上のtotalRNAが必要であるため、入手量が限られる希少サンプル等の研究は非常に困難です。このサービスで使用するアレイは、3μgのtotalRNAしか必要としないため、MeRIP-Seqを大幅に下回るスタートサンプルで解析が可能です。
| サンプル条件 |
エピトランスクリプトームマイクロアレイ |
MeRIP-Seq |
| スタートサンプル量 |
≧3μg total RNA |
≧120μg total RNA |
| mRNAおよびrRNAの除去 |
不要 |
必要 |
| インタクトRNA |
不要 |
必要 |
【マイクロアレイによる各転写産物の修飾率の定量化】
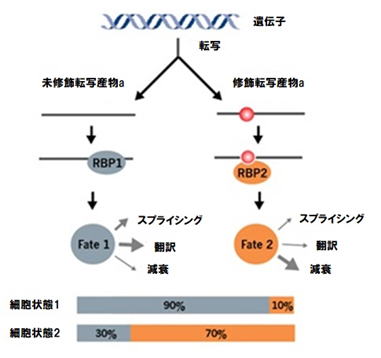
同じRNA転写産物の修飾画分と未修飾画分は、構造やそれらに結合するリードタンパク質のみの違いによって、異なった運命をたどることができます。重要なことに、変化した転写産物の割合は、その後の機能的な結果に非常に大きく関連します(
fig.1)。
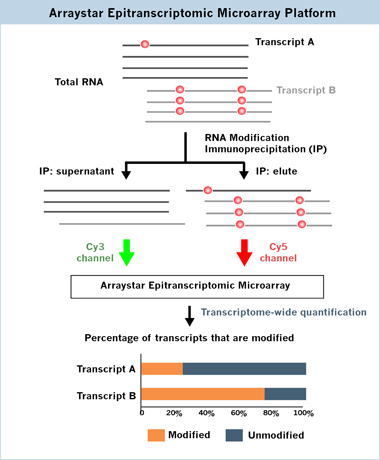
MeRIP-seq(すなわち m6A-seq)などの現在のRNA修飾プロファイリング法では、修飾部位のマッピングは可能ですが、特定の転写産物の修飾RNAと未修飾RNAの相対的な割合を定量化することはできません。本サービスでは、マイクロアレイ上で2つの蛍光チャネルを使用し、各転写産物の修飾度合を測定することで、修飾された転写産物の割合を決定することが可能です(
fig.2)。また、それと同時にどの遺伝子転写物が修飾されているか、異なる条件間の修飾レベルをプロファイリングすることが可能です。
(
fig.1)
変化する修飾化学量論は、同じRNA転写物から機能的多様性を生み出します。修飾RNA(転写産物a)の割合は、ある細胞条件下(例えば細胞状態1)では非常に低くなり得るが、別の細胞条件下(例えば細胞状態2)では高く変化する場合があります。RNAの構造変化や修飾リードタンパク質の直接動作に引き起こすことで、修飾された(転写産物a)は、例えばタンパク質翻訳から、増加したRNAの崩壊まで、異なる運命を獲得します。
(
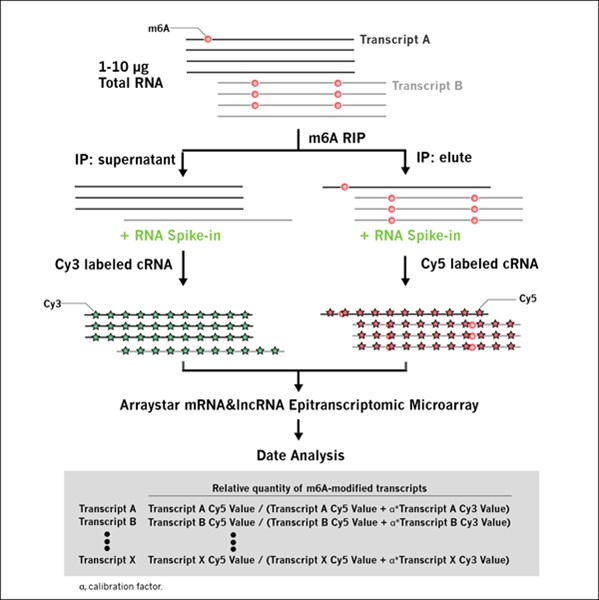
fig.2)免疫沈降法による m6A 濃縮法
エピトランスクリプトームマイクロアレイは、Cy5チャンネルで免疫沈降した修飾RNAと、Cy3チャンネルで上清の未修飾RNAをそれぞれ検出し、各転写産物の修飾および未修飾の割合を測定します。これらスプライスされた転写物アイソフォームは、転写物特異的なアレイプローブによって特異的かつ明確に検出します。
(
fig.3)GST-YTH 法による m6A 濃縮法
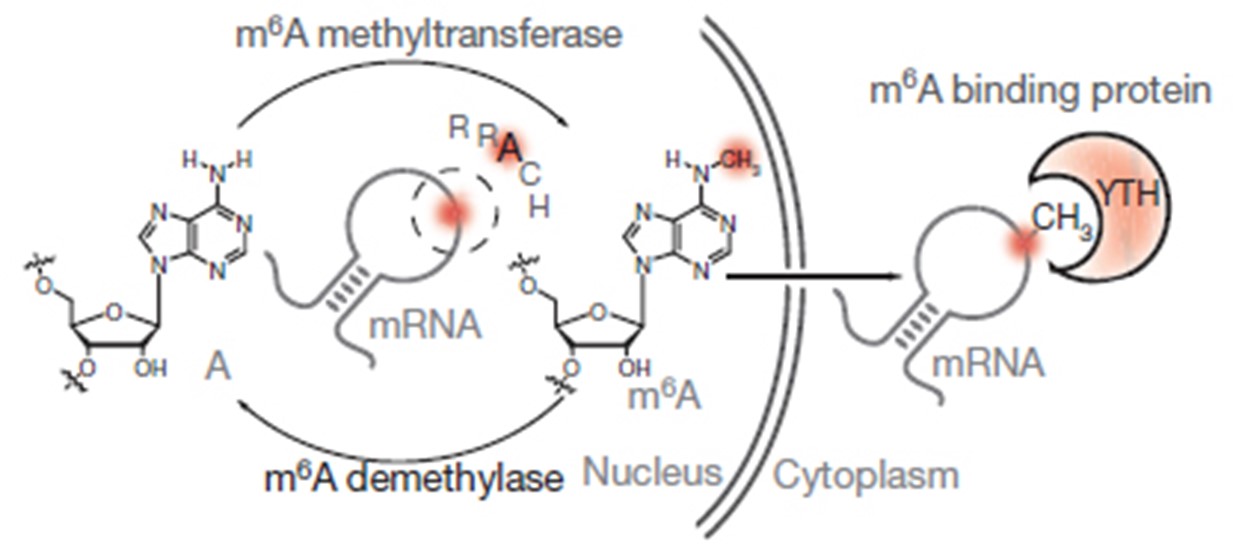
GST-YTH は、YTH-DF2 m6A リーダードメイン(a.a385-579)と m6A RNA 濃縮用 GST タグの組換え融合タンパク質である。YTH は進化的に保存された構造ドメインであり、コンセンサス RRACH モチーフ内の m6A を選択的に「読み取り」、結合します[1]。構造的には、YTH ドメインには 2~3 個のトリプトファン残基があり、m6A に対して芳香族ケージと結合ポケットを形成し、さらに m6A の前後のヌクレオチドとも相互作用するため、RRACH モチーフに優先的な配列を与えます[2-4]。すなわち、GST-YTH は m6A 構造および RRACH 配列モチーフに依存した方法で m6A を含む RNA と結合し[1]、その両方が m6Am や他の類似の RNA 修飾とは異なります。したがって、GST-YTH の pull down 法は、m6A 抗体 MeRIP による他の構造的に類似した RNA 修飾[5]、特に m6Am[6]との交差反応性を示すことなく、m6A に対して高度に特異的です。
・GST-YTHに関する参考文献
1. Wang X et al: N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505(7481):117-120.[PMID: 24284625]
2. Luo S, Tong L: Molecular basis for the recognition of methylated adenines in RNA by the eukaryotic YTH domain. Proc Natl Acad Sci U S A 2014, 111(38):13834-13839.[PMID: 25201973]
3. Theler D et al: Solution structure of the YTH domain in complex with N6-methyladenosine RNA: a reader of methylated RNA. Nucleic Acids Res 2014, 42(22):13911-13919.[PMID: 25389274]
4. Xu C et al: Structural basis for selective binding of m6A RNA by the YTHDC1 YTH domain. Nat Chem Biol 2014, 10(11):927-929.[PMID: 25242552]
5. Linder B et al: Single-nucleotide-resolution mapping of m6A and m6Am throughout the transcriptome. Nat Methods 2015, 12(8):767-772.[PMID: 26121403]
6.
メーカーの製品情報サイト
【coding および non-coding なエピトランスクリプトームのカバレッジ】
本サービスでは、以下の2種類のマイクロアレイからご選択いただき、解析を行うことが可能です。マイクロアレイのスライド構成等は以下の「マイクロアレイデータベース」をご参照ください。
Arraystar mRNA&lncRNA Epitranscriptomic Microarray
mRNA、lncRNA および中サイズの non-coding RNA クラス(pre-miRNA、pri-miRNA、snoRNA、snRNA)をカバー
- Arraystar circRNA Epitranscriptomic Microarray
4サンプル以上かつ2試験以上で観察された発現の信頼性が高い環状RNAをカバー
MeRIP-seq では難しいRNA(lncRNAやcircRNAなど)でも高い感度と精度を発揮します。サンプル中のlncRNA/circRNAジャンクション部位の存在量は非常に低く、シークエンシングによって正確に定量することはできません。IPエンリッチなlncRNA/circRNAジャンクションについても同様です。
lncRNA プロファイリングに対する RNA-seqの制限について
Circular RNA 発現プロファイリングに RNA-seq よりもマイクロアレイを使用する理由について
【転写産物特異的な RNA 修飾レベル】
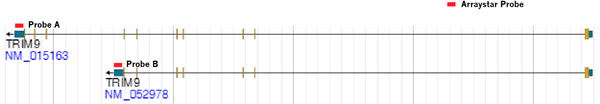
スプライシングされた転写物アイソフォームは、組織特異的な発現と、異なる生化学的機能を持っています。例えば、TRIM9の長いアイソフォーム(NM_015163)ではなく短いアイソフォーム(NM_052978)は、ウイルス感染に応答して、炎症性サイトカインの生産を選択的に阻害します。このことからも、修飾された転写物アイソフォームの比率の変化が生化学的機能や疾患に関連していることが分かります。
本サービスに使用するマイクロアレイは、特定のエクソンまたはスプライスジャンクションプローブを採用しており、個々の転写物アイソフォームのRNA修飾を正確にプロファイリングし、エピトランスクリプトミクスをより詳細に定義することができます(
fig.3)。
Arraystar Epitranscriptpmic Microarray は、転写物特異的なプローブAとプローブBを使用して、TRIM9の長い(NM_015163)および短い(NM_052978)転写物アイソフォームのRNA修飾を明確に検出します。
マイクロアレイデータベース
本サービスは、解析内容に合わせて、mRNA & lncRNA解析用のアレイと、circRNA解析用のアレイの2タイプからご選択いただけます。
【mRNA & lncRNA エピトランスクリプトームマイクロアレイ】
| プローブ総数 |
60,613(Human)、60,773(Mouse)、40,991(Rat) |
| プローブ長 |
60nt |
| プローブサイト |
mRNA & lncRNA:転写物の全長に沿った特異的エクソンまたはスプライスジャンクション配列。
中サイズncRNA:ncRNA内のユニークな配列領域。 |
| プローブ特異性 |
転写物特異的 |
| 標識法 |
cRNAは、少量の分解RNAでも3’バイアスなしで全長に沿って標識。 |
| タンパク質コードmRNA数 |
44,122(Human)、48,161(Mouse)、27,770(Rat) |
| lncRNA数 |
12,496(Human)、8,393(Mouse)、10,582(Rat) |
| 中サイズncRNA数 |
Human:1,366(pre-miRNA)、1,642(pri-miRNA)、19(snRNA)、786(snoRNA)
Mouse:701(pre-miRNA)、957(pri-miRNA)、1,229(snRNA)、1,200(snoRNA)
at:432(pre-miRNA)、449(pri-miRNA)、155(snRNA)、1,469(snoRNA) |
| mRNAソース |
Human:Refseq, UCSC, GENECODE, FANTOM5 CAT[2-6]
Mouse:Refseq, UCSC, GENECODE[2-3]
Rat:Refseq, Ensembl 92[1, 5] |
| lncRNAソース |
- Arraystar lncRNA コレクションパイプライン
2018年までのすべてのデータベースおよび文献からのlncRNA
各lncRNA遺伝子の転写物に割り当てられた「Canonical」または「longest」の優先順位。
- 外部データベース(2018年現在)
Human:Refseq, UCSC, GENCODE, FANTOM5 CAT, LncRNAdb, RNAdb, NRED [2-8]
Mouse:Refseq, UCSC, GENCODE, GenBank, lncRNAdb, NRED[2-7]
Rat:Ensembl 92[1], Refseq [5]
- 文献
Human: Scientific publications up to 2018[9-45]
Mouse: lincRNA catalogs[8-11], T-UCRs[12], Evolutionary constrained LncRNAs[13], lncRNA publications up to 2018[14-22]
Rat:T-UCRs[6-9]
|
| 中サイズncRNAソース |
HumanおよびMouse:GENECODE, miRBase[1-2]
Rat:GENECODE, miRBase, GtRNADb[2-3] |
| アレイフォーマット |
8×60K |
リファレンスリスト(PDF):Human、Mouse、Rat
*生物種をクリックするとダウンロードできます。
【circRNA エピトランスクリプトームマイクロアレイ】
| プローブ総数 |
13,617(Human)、14,236(Mouse)、14,145(Rat) |
| プローブ長 |
60nt |
| プローブサイト |
環状ジャンクション |
| プローブ特異性 |
転写物特異的 |
| 標識法 |
環状RNAの特異的かつ効率的な標識を確実にするためのRNase Rサンプル前処理と組み合わせたランダムプライマー標識。 |
| circRNA数 |
13,617(Human)、14,236(Mouse)、14,145(Rat) |
| circRNAソース |
- 外部データベース(2018年現在)
circbase, CircNet, circRNADb[1-3]
- 文献
Human:[4-10]
Mouse:scientific publications[4-5]
Rat:scientific publications[4-5]
|
| アレイフォーマット |
8×15K |
リファレンスリスト(PDF):Human、Mouse、Rat
*生物種をクリックするとダウンロードできます。
解析の流れ
本サービスは、MeRIP、cRNA標識、アレイ実験、アノテーションおよびデータ解析まで、フルパッケージとなっています。
【mRNA & lncRNA 解析ワークフロー】(
fig.4)
- RNAサンプルの受け取り
- RNAのQCチェック
- 免疫沈降(MeRIP)
- cRNA合成、および2色標識(IP-RNAをCy5標識、およびCy3上清RNAをCy3標識)
- アレイハイブリダイゼーション、洗浄、およびスキャニング
- データ抽出、アノテーション、解析および要約
【circRNA解析ワークフロー】(
fig.5)
- RNAサンプルの受け取り
- RNAのQCチェック
- 免疫沈降(MeRIP)
- RNase RによるリニアRNAの除去(rRNA、lncRNA、mRNAなど)
- cRNA合成、および2色標識(RIP-RNAをCy5標識、およびCy3上清RNAをCy3標識)
- アレイハイブリダイゼーション、洗浄、およびスキャニング
- データ抽出、アノテーション、解析および要約
【バイオインフォマティクス解析】
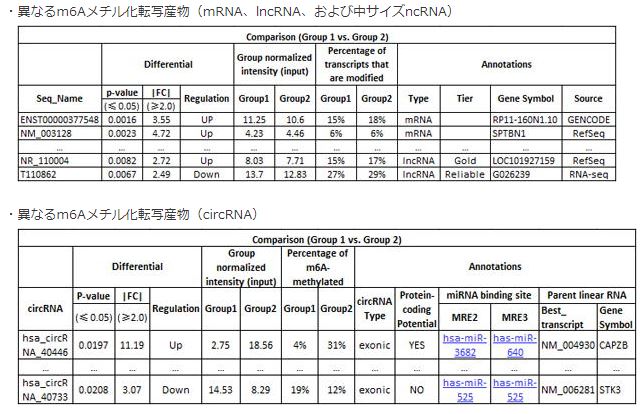
パッケージ内では、以下のような内容の解析が行われます。
- 異なるm6Aメチル化転写産物(mRNA、lncRNA、および中サイズncRNA)(fig.6)
- 異なるm6Aメチル化転写産物(circRNA)(fig.6)
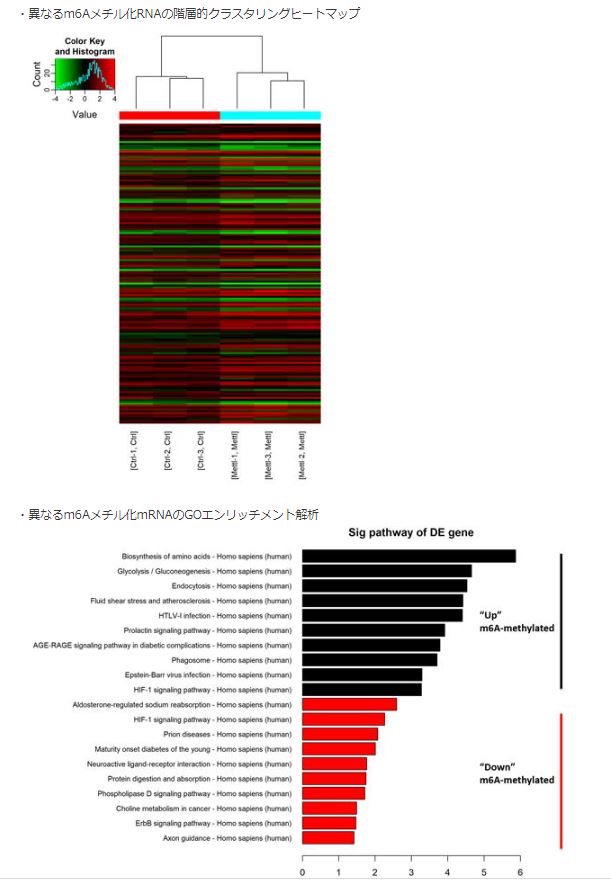
- 異なるm6Aメチル化RNAの階層的クラスタリングヒートマップ(fig.7)
- 異なるm6Aメチル化mRNAのGOエンリッチメント解析(fig.7)
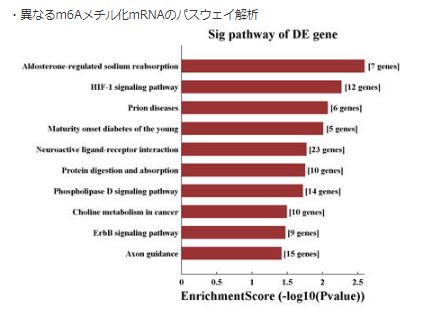
- 異なるm6Aメチル化mRNAのパスウェイ解析(fig.8)
参考文献
- Gilbert WV, Bell TA, Schaening C: messenger RNA modifications: Form, distribution, and function. Science 2016, 352(6292):1408-1412.[PMID: 273113037]
- Yang Y et al: Extensive translation of circular RNAs driven by N(6)-methyladenosine. Cell Res 2017, 27(5):626-641.[PMID:28281539]
- Alarcon CR et al: HNRNPA2B1 is a Mediator of m(6)A-Dependent Nuclear RNA Processing Events. Cell 2015, 162(6):1299-1308.[PMID:26321680]
- Lewis CJ, Pan T, Kalsotra A: RNA modifications and structures cooperate to guide RNA-protein interactions. Nat Rev Mol Cell Biol 2017, 18(3):202-210.[PMID:28144031]
- Qin Y et al: TRIM9 short isoform preferentially promotes DNA and RNA virus-induced production of type I interfe*** by recruiting GSK3beta to TBK1. Cell Res 2016, 26(5):613-628.[PMID: 26915459]